コラム
ColumnPhenoVation社 クロロフィル蛍光測定法:OJIPプロトコル – 第2部:JIPテストのパラメータとその活用
クロロフィル蛍光測定法:OJIPプロトコル – 第2部:JIPテストのパラメータとその活用
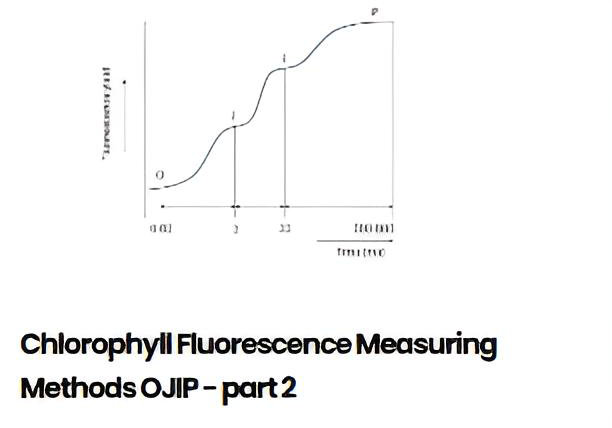
OJIPは、クロロフィル蛍光ダイナミクス、特に飽和光パルス照射時における初期の急速な蛍光上昇(ライズ)プロセスを詳細に調査するために用いられる測定手法です。この上昇プロセスを解析することで、光合成機構内におけるエネルギーフラックス(流束)や電子伝達に関する極めて重要な情報を得ることができます。
このOJIPブログシリーズの第1部では、どのような生理学的プロセスがこの特徴的な蛍光誘導曲線の立ち上がりを引き起こすのかを解説しました。今回の第2部では、OJIPの一連の過渡変化から導き出される様々なパラメータ(JIPテスト)に焦点を当てます。これらのパラメータを使用することで、前回解説した各生理プロセスを数値化し、曲線の形状を測定可能な生理学的指標へと変換することができます。
1. パラメータの概要と分類
JIPテストの各パラメータは、光合成システム内におけるエネルギーの経路に焦点を当てています。すなわち、光エネルギーがアンテナ色素に吸収された瞬間からその運命を追跡し、それが光合成の駆動に使用されたのか、あるいは熱や蛍光として散逸したのかを示します。これらのパラメータは大きく分けて、「エネルギーフラックス」「量子収率」「効率」の3つのカテゴリに分類できます。
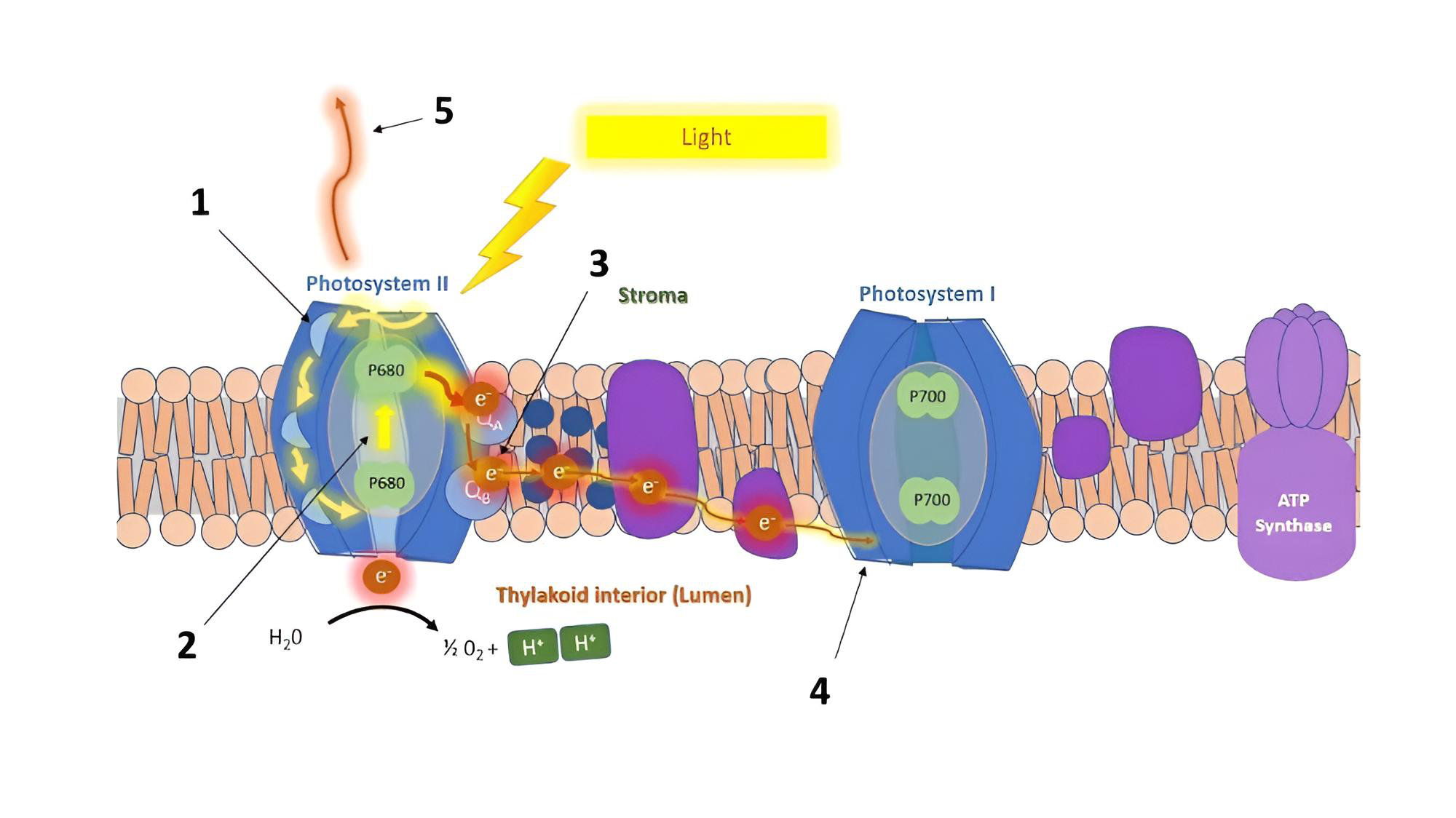
1.1. エネルギーフラックス(Energy fluxes)
フラッシュ中の各反応中心(RC)、または被照射面積(CS)あたりを通過するエネルギーの流れの量を記述するパラメータです。
- (1) 吸収フラックス(Absorption flux / ABS): アンテナ色素によって吸収され、励起クロロフィルを生成した光子(フォトン)の量。
- (2) 捕獲フラックス(Trapping flux / TR): 励起クロロフィルから反応中心へと誘導され、電子伝達鎖へと変換された($Q_A$の還元に回された)エネルギー。
- (3) 電子伝達フラックス(Electron transport flux / ET): $Q_A$の先へと流れていく電子伝達の量。
- (4) 還元フラックス(Reduction flux / RE): 光化学系I(PSI)側において、最終的な電子受容体を還元するフラックス。
- (5) 散逸フラックス(Dissipation flux / DI): 光合成に使用されず、熱や蛍光として能動的・受動的に失われたエネルギーの量。
1.2. 量子収率(Quantum yields)および効率(Efficiencies)
エネルギーフラックスと並行して、光化学系II(PSII)によって吸収されたエネルギーが、特定の光化学的イベントにどれだけの割合で割り振られたかを計算できます。この割合を「量子収率」と呼びます。また、捕獲されたエネルギーをベースとした各イベントの「効率」も算出可能です。
- QAを還元する一次光化学反応の量子収率(Φp0): 一般的にFv/Fmとして表現されるパラメータです。
- QA → QB → PQ プールの電子輸送の量子収率 (φE₀)
- 最終的なPSI電子受容体への電子伝達の量子収率(ΦR0)
- 捕獲された励起エネルギーが $Q_A$ を越えて電子伝達鎖の先へと電子を移動させる効率(ΨE₀)
- PSIに到達した電子が、最終受容体(FA/FB、フェレドキシン、またはNADP⁺ )を還元する効率(δR₀)
これらのパラメータは、光合成機構の機能的状態に関する深い洞察を提供するため、様々な資材・化合物の作用機序(モード・オブ・アクション)の解明や、環境ストレスの影響を精緻に調査する研究において極めて価値の高い指標となります。
そして、これらの複数のプロセスを統合し、植物のストレスに対して極めて高い感度を示す代表的なパラメータがパフォーマンスインデックス(Performance Index: PI)です。
1.3. パフォーマンスインデックス(Performance Index / PI)
一般的に光合成効率の指標として多用される $F_v/F_m$ は、ストレスの初期段階や軽微なストレス応答に対して、必ずしも十分な感受性(感度)を持たない場合があります。そこで導入されたのがパフォーマンスインデックス(PI)です。PIは、光合成プロセスの複数のコンポーネント(エネルギーの吸収、捕獲、および電子伝達)を単一の数値に統合した指標です。植物生理学者によって、生物的・非生物的ストレスの影響を包括的に評価するために広く利用されているほか、乾燥ストレスや塩害ストレス下での「耐性系統の迅速なスクリーニング」を行う育種プログラムにおいても非常に強力なパラメータとして機能します。PIは、OJIPから導き出されるパラメータの中で最も繊細な(感度の高い)指標のひとつと考えられています。
パフォーマンスインデックスには、PI_ABSとPI_TOTALの2つのバリエーションが存在します。
PI_ABSは、以下の3つの要素を1つの値に統合しています:
- 活性な反応中心の密度(The density of active reaction centres)
- エネルギー捕獲の効率(吸収された光エネルギーがQAを閉じる確率)
- QAを越えた先の電子伝達の効率
PI_TOTALは、上記のPI_ABSの3要素に、さらに4つ目の要素を追加したものです:
- 最終的なPSI電子受容体まで電子が伝達される効率
PIは光合成電子伝達鎖における複数のステップを包括しているため、どこか1つのコンポーネントにでも障害(ストレスの影響)が発生すれば、それが全体のPI値に敏感に反映されます。そのため、個々の単一パラメータを単独で確認するよりも、遥かに高精度に異常をキャッチすることができます。
2. 文献レビュー:各種ストレスがパラメータに与える影響
JIPテストのパラメータが実際の研究でどのように活用されているかのオーバービューを提供するため、私たちは各種ストレス(塩害ストレス、乾燥ストレス、熱ストレス、低温ストレス、強光ストレス)が各パラメータにどのような影響を及ぼすかを網羅した学術文献の調査を行いました。
一般的な傾向として、特定のJIPパラメータは様々な研究にわたりストレスに対して一貫した応答を示しますが、その変化の方向性(数値の上昇・下落)は一様ではありません。特定の種や栽培環境下では数値が上昇する一方で、別の条件下では減少することもあります。この詳細な挙動パターンは、植物の種や品種、およびストレスの深刻度(進行度)に依存します。調査結果のサマリーをまとめたデータテーブルは、以下のリンクよりご確認いただけます。
3. OJIPパラメータの可視化とデータ解釈の手法
JIPテストから得られる膨大な数のパラメータは、一見するとデータ解釈の難易度を高く感じさせるかもしれません。しかし、個々のパラメータが光合成機構の「異なるコンポーネント」を個別に反映しているため、データセット全体を網羅的に確認することは研究者にとって極めて有意義です。電子伝達鎖のある一部分で起きた変化が、別の部分には現れないケースもあるため、複数のパラメータを多角的に検査することで、変化やストレス応答が「具体的にどの部位で発生しているのか」をピンポイントに特定できます。
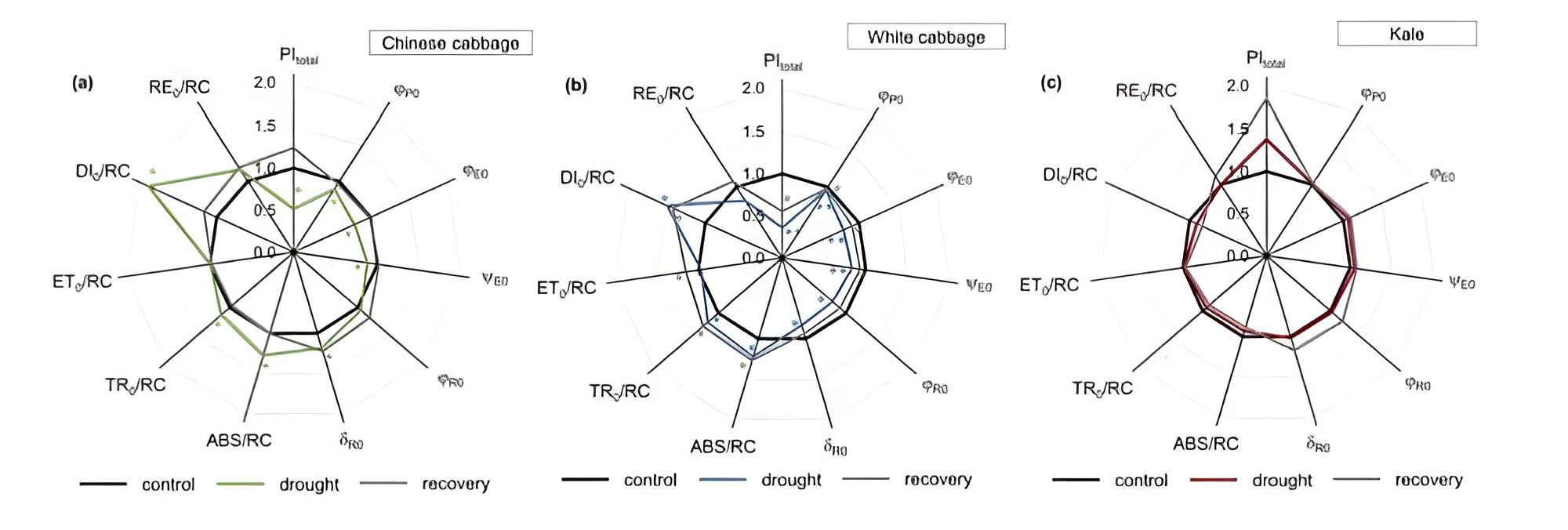
この包括的なデータセットを効果的に視覚化する優れた手法として、スパイダープロット(レーダーチャート / 図2)がよく用いられます。多くのパラメータを同一グラフ上に同時に表示できるため、パターンの特定、各処理区間の比較、あるいはストレスに起因する標準値からの逸脱を直感的に捉えることが容易になります。
また、一部の研究論文では、被照射面積あたりの各エネルギーフラックスの挙動を、以下のような「葉モデル(Leaf Model / 図3)」という模式図を用いて可視化しています。このスキーマ(回路図)では、矢印の太さが各エネルギーフラックスの大きさ(流量)を表しています。黄色が吸収フラックス(ABS)、濃い青色が電子伝達フラックス(ET)、水色が捕獲フラックス(TR)、そして赤色が散逸フラックス(DI)を示します。中央の正方形の内部にある黒い点は、不活性化した(機能停止した)反応中心の数を表しています。
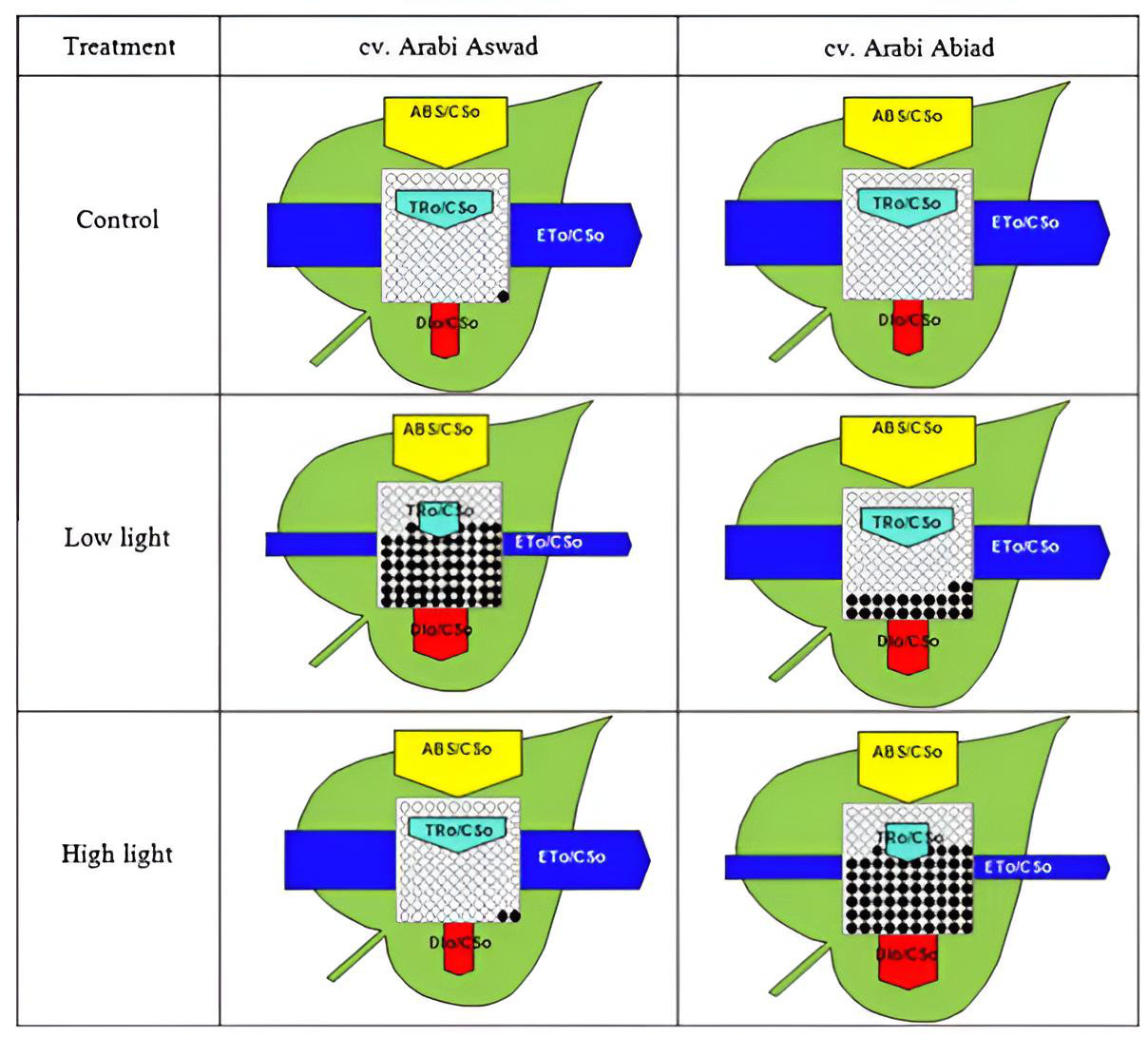
このタイプのモデルは、光合成機構のリアルタイムな状態を1枚のインフォグラフィックスとして要約できるため、非常に示唆に富んでいます。上記の例では、強光条件下と弱光条件下における大麦(バーレイ)の2つの品種を比較しています。フラックスのパターンから、「Arabi Aswad」という品種(左)の方が、より大きな電子伝達フラックスを維持し、散逸フラックスが少なく、活性な反応中心を多く保てていることから、強光ストレスの処理能力に長けていることが明瞭に分かります。一方、「Arabi Abiad」という品種(右)は、弱光環境により適応していることが読み取れます。
このモデルは、反応中心(RC)あたりのフラックスを示す「膜モデル(Membrane Model / 図4)」として表現することも可能です。以下にその一例を示します。
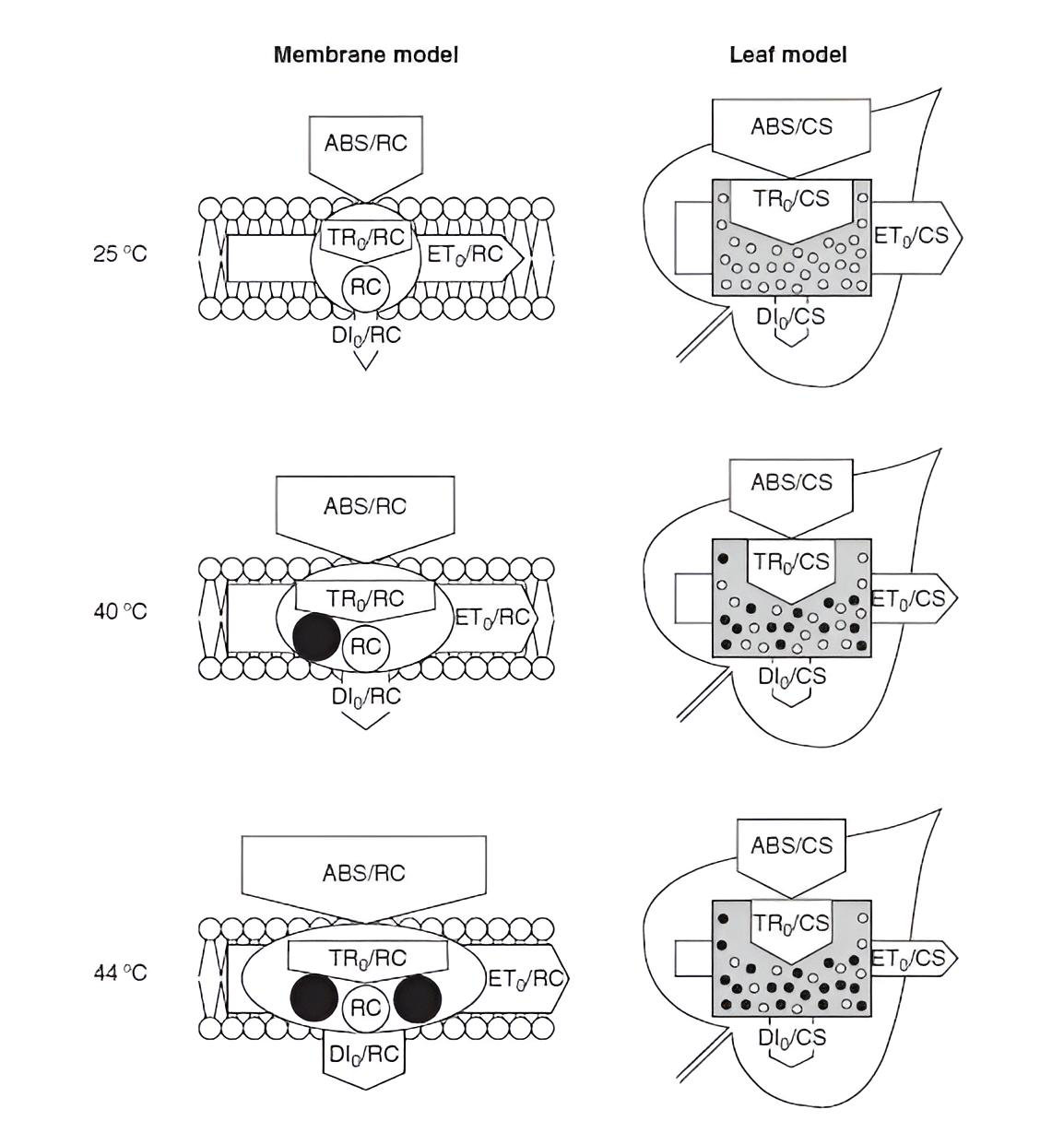
一例として、熱ストレス(加熱)が植物に与える影響について、ストラッサーは膜モデル(反応中心あたり)の変化を以下のように記述しています:
- 「一部の反応中心(RC)が熱阻害によって不活性化するため、生存してアクティブに残っている反応中心あたりの吸収フラックス(ABS/RC)が相対的に増大する」
- 「不活性化した反応中心からの熱放散が高まるため、活性な反応中心あたりの総散逸フラックス(DI/RC)の比率が増大する」
- 「暗反応の熱活性化(酵素反応の加速)に伴い、活性な反応中心あたりの電子伝達フラックス(ET/RC)が増大する」
一方で、面積ベースの葉モデル(被照射断面あたり)における熱の影響については以下のように説明されています:
- 「反応中心複合体の不活性化に伴い、励起断面積あたりの電子伝達フラックス(ET/CS)が減少する」
- 「活性な反応中心の密度(RC/CS:白抜きの円で示される)が減少する」
- 「励起断面積あたりのエネルギー散逸フラックス(DI/CS)が増大する」
- 「励起断面積あたりの総吸収エネルギーフラックス(ABS/CS)が減少する」
このように、JIPテストのパラメータを2つのモデル(面単位・点単位)で捉え直すことで、植物の内部で起きている複雑な光合成のダイナミクスを直感的に理解し、栽培環境の最適化や植物の活力評価へと確信を持って応用することができます。
4. 参考文献(References)
- Strasser, R.J., Srivastava, A. and Govindjee (1995). Polyphasic chlorophyll a fluorescence transient in plants and cyanobacteria. Photochemistry and Photobiology 61: 32-42.
- Strasser, R.J., Srivastava, A. and Tsimilli-Michael, M. (2000). The fluorescence transient as a tool to characterize and screen photosynthetic samples. In: Yunus, M. (Ed.) Probing Photosynthesis: Mechanisms, Regulation and Adaptation. Taylor and Francis, London: 445-483.
- Strasser, R.J., Tsimilli-Michael, M. and Srivastava, A. (2004). Analysis of the Chlorophyll a fluorescence transient. In: Papageorgiou, G.C. and Govindjee (Eds.) Chlorophyll a fluorescence: a signature of photosynthesis. Springer, Dordrecht: 321-362.
- Lotfi, R. et al. (2018). Effects of humic acid on photosynthetic efficiency of rapeseed plants growing under different watering conditions. Photosynthetica 56(3): 962–970.
- Zushi, K. et al. (2012). Chlorophyll a fluorescence OJIP transient as a tool to characterize and evaluate response to heat and chilling stress in tomato leaf and fruit. Scientia Horticulturae 148: 39–46.
- Kalaji, H. M. et al. (2018). Prompt chlorophyll fluorescence as a tool for crop phenotyping: an example of barley landraces exposed to various abiotic stress factors. Photosynthetica 56: 953–961.
- Chen, X. et al. (2021). Ascorbic acid‑induced photosynthetic adaptability of processing tomatoes to salt stress probed by fast OJIP fluorescence rise. Frontiers in Plant Science 12: 594400.
- Stefanov, D. et al. (2011). Screening for heat tolerance in common bean (Phaseolus vulgaris L.) lines and cultivars using JIP‑test. Scientia Horticulturae 128(1): 1–6.
- Jedmowski, C., and W. Brüggemann. (2015). Imaging of fast chlorophyll fluorescence induction curve (OJIP) parameters, applied in a screening study with wild barley (Hordeum spontaneum) genotypes under heat stress. Journal of Photochemistry and Photobiology B: Biology 151: 153–160.
- Kalaji, H. M. et al. (2014). Identification of nutrient deficiency in maize and tomato plants by in vivo chlorophyll a fluorescence measurements. Plant Physiology and Biochemistry 81: 16–25.
- Mehta, P. et al. (2010). Characterization of photosystem II heterogeneity in response to high salt stress in wheat leaves (Triticum aestivum). Photosynthesis Research 105(3): 249–255.
- Jedmowski, C. et al. (2013). Reactions of Egyptian landraces of Hordeum vulgare and Sorghum bicolor to drought stress, evaluated by the OJIP fluorescence transient analysis. Acta Physiologiae Plantarum 35: 345–354.
- Ren, J. et al. (2025). Differential photosynthetic responses to drought stress in peanut varieties: insights from transcriptome profiling and JIP‑Test analysis. BMC Plant Biology 25: 957.
- Mihaljević, I. et al. (2024). Assessment of photosynthetic capacity of two blackberry cultivars subjected to salt stress by the JIP fluorescence test. Journal of Berry Research 14(1): 1–13.
- Rastogi, A. et al. (2020). JIP‑test as a tool to identify salinity tolerance in sweet sorghum genotypes. Photosynthetica 58: 518–528.
- Ranjbarfordoei, A. et al. (2006). Chlorophyll fluorescence performance of sweet almond (Prunus dulcis) in response to salinity stress induced by NaCl. Photosynthetica 44(4): 513–522.
- Jafarinia, M., and M. Shariati. (2012). Effects of salt stress on photosystem II of canola plant (Brassica napus L.) probed by chlorophyll a fluorescence measurements. Iranian Journal of Science and Technology (Sciences) 36(1): 71–76.
- Zushi, K., and N. Matsuzoe. (2017). Using of chlorophyll a fluorescence OJIP transients for sensing salt stress in the leaves and fruits of tomato. Scientia Horticulturae 219: 216–221.
- Giorio, P., and M. H. Sellami. (2021). Polyphasic OKJIP chlorophyll a fluorescence transient in a landrace and a commercial cultivar of sweet pepper (Capsicum annuum L.) under long‑term salt stress. Plants 10(5): 887.
- Akhter, M. S. et al. (2021). Influence of salinity stress on PSII in barley (Hordeum vulgare L.) genotypes, probed by chlorophyll‑a fluorescence. Journal of King Saud University – Science 33(1): 101239.
- Zhang, T. et al. (2010). Salt stress induces a decrease in excitation energy transfer from phycobilisomes to photosystem II but an increase to photosystem I in the cyanobacterium Spirulina platensis. Journal of Plant Physiology 167(12): 951–958.
- Weng, H. et al. (2023). High‑throughput phenotyping salt tolerance in JUNCAOs by combining prompt chlorophyll a fluorescence with hyperspectral spectroscopy. Plant Science 330: 111660.
- Chen, X. et al. (2025). Melatonin alleviates photosynthetic injury in tomato seedlings subjected to salt stress via OJIP chlorophyll fluorescence kinetics. Plants 14(5): 824.
- Antunović Dunić, J. et al. (2023). Comparative analysis of primary photosynthetic reactions assessed by OJIP kinetics in three Brassica crops after drought and recovery. Applied Sciences 13(5): 3078.
- Zhang, R. H. et al. (2015). Photosynthetic performance of maize hybrids to drought stress. Russian Journal of Plant Physiology 62: 788–796.
- Meng, L.‑L. et al. (2016). Effects of drought stress on fluorescence characteristics of photosystem II in leaves of Plectranthus scutellarioides. Photosynthetica 54(3): 414–421.
- Zhou, R. et al. (2019). Drought‑induced changes in photosynthetic electron transport in maize probed by prompt fluorescence, delayed fluorescence, P700 and cyclic electron flow signals. Environmental and Experimental Botany 158: 51–62.
- Liu, J. et al. (2018). Effects of drought stress on the photosynthesis in maize. Russian Journal of Plant Physiology 65: 849.
- Zhang, K. et al. (2018). Effects of short‑term heat stress on PSII and subsequent recovery for senescent leaves of Vitis vinifera L. cv. Red Globe. Journal of Integrative Agriculture 17(12): 2683–2693.
- Zhang, L. et al. (2017). The alleviation of heat damage to photosystem II and enzymatic antioxidants by exogenous spermidine in tall fescue. Frontiers in Plant Science 8: 1747.
- Fan, Q., and D. Jespersen. (2023). Assessing heat tolerance in creeping bentgrass lines based on physiological responses. Plants 12(1): 41.
- Tan, W. et al. (2011). Photosynthesis is improved by exogenous calcium in heat‑stressed tobacco plants. Journal of Plant Physiology 168: 2063–2071.
- Arslan, Ö. (2023). The role of heat acclimation in thermotolerance of chickpea cultivars: Changes in photochemical and biochemical responses. Life 13(1): 233.
- Mihaljević, I. et al. (2025). Characterization of heat tolerance in two apple rootstocks using chlorophyll fluorescence as a screening method. Agronomy 15(6): 1442.
- Kalaji, H. M. et al. (2012). Fluorescence parameters as early indicators of light stress in barley. Journal of Photochemistry and Photobiology B: Biology 112: 1–6.
- Lee, J. H. et al. (2021). Chlorophyll fluorescence and growth evaluation of ornamental foliage plants in response to light intensity levels under continuous lighting conditions. Flower Research Journal 29(4): 320–322.
- Bayat, L. et al. (2018). Effects of growth under different light spectra on the subsequent high light tolerance in rose plants. AoB Plants 10(5): ply052.
- Ceusters, N. et al. (2019). Performance Index and PSII connectivity under drought and contrasting light regimes in the CAM orchid Phalaenopsis. Frontiers in Plant Science 10: 1012.
- Faseela, P., and J. T. Puthur. (2016). Chlorophyll a fluorescence changes in response to short‑ and long‑term high‑light stress in rice seedlings. Photosynthetica 54(3): 549–558.
- Faseela, P., and J. T. Puthur. (2018). The imprints of the high light and UV‑B stresses in Oryza sativa L. ‘Kanchana’ seedlings are differentially modulated. Journal of Photochemistry and Photobiology B: Biology 178: 551–559.
- Mlinarić, S. et al. (2017). Differential accumulation of photosynthetic proteins regulates diurnal photochemical adjustments of PSII in common fig (Ficus carica L.) leaves. Journal of Plant Physiology 209: 1–10.
- Lu, T. et al. (2019). Improving plant growth and alleviating photosynthetic inhibition and oxidative stress from low‑light stress with exogenous GR24 in tomato (Solanum lycopersicum L.) seedlings. Frontiers in Plant Science 10: 490.
- Li, L. et al. (2013). Assimilative branches and leaves of the desert plant Alhagi sparsifolia Shap. possess a different adaptation mechanism to shade. Plant Physiology and Biochemistry 74: 239–245.
- Liang, Y. et al. (2007). Responses of Jatropha curcas seedlings to cold stress: photosynthesis‑related proteins and chlorophyll fluorescence characteristics. Physiologia Plantarum 131(3): 508–517.
- Huang, X. et al. (2017). ABA is involved in regulation of cold stress response in Bermudagrass. Frontiers in Plant Science 8: 1613.
- Mazur, M. et al. (2024). Crop‑specific responses to cold stress and priming: Insights from chlorophyll fluorescence and spectral reflectance analysis in maize and soybean. Plants 13(9): 1204.
- Refan, J. et al. (2015). Alleviation of cold damage to photosystem II and metabolisms by melatonin in Bermudagrass. Frontiers in Plant Science 6: 925.
- Li, J. et al. (2023). Magnesium application improves the morphology, nutrient uptake, photosynthetic traits, and quality of tobacco (Nicotiana tabacum L.) under cold stress. Frontiers in Plant Science 14: 1078128.
- Sun, L. et al. (2018). Cold priming induced tolerance to subsequent low temperature stress is enhanced by melatonin application during recovery in wheat. Molecules 23(5): 1091.
- Tsimilli-Michael, M. (2020). Revisiting JIP-test: An educative review on concepts, assumptions, approximations, definitions and terminology. Photosynthetica 58: 275-292.





